Ji Hye Kim1,
Min Young Kim1,2 ![]()
For correspondence:- Min Young Kim Email: jeffmkim@jejunu.ac.kr Tel:+82647543349
Received: 19 February 2016 Accepted: 22 May 2016 Published: 28 June 2016
Citation: Kim J, Kim M. Apoptotic properties of Citrus sudachi Hort, ex Shirai (Rutaceae) extract on human A549 and HepG2 cancer cells. Trop J Pharm Res 2016; 15(6):1167-1174 doi: 10.4314/tjpr.v15i6.8
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate whether Citrus sudachi harvested at two stages of maturity can induce toxicity in a cell-specific manner and to determine the possible mechanisms of Citrus sudachi-induced cytotoxic responses in two types of cancer cells (human lung adenocarcinoma A549 and hepatocellular carcinoma HepG2 cells) and two normal cell lines (lung 16HBE140- and liver CHANG cells).
Methods: 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) and annexin V/propidium iodidle assay were used to test the antiproliferative activity and apoptosis of methanol extract of Citrus sudachi, respectively. Griess reaction and reverse transcriptase-polymerase chain reaction (RT-PCR) were carried out to evaluate nitric oxide (NO•) production and the mRNA levels of inhibitors of apoptosis (IAP).
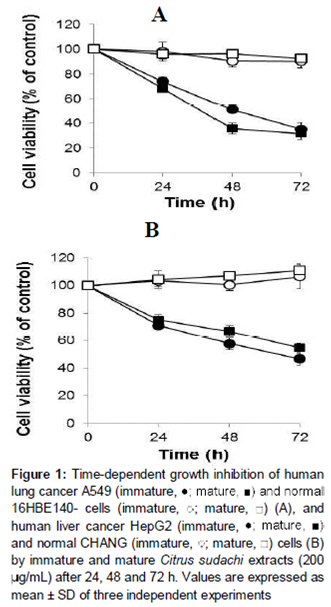
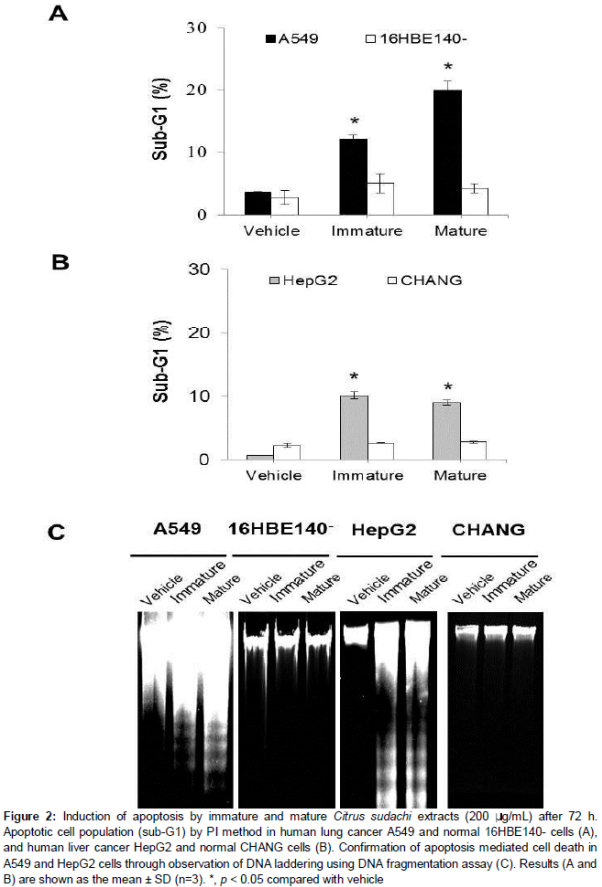
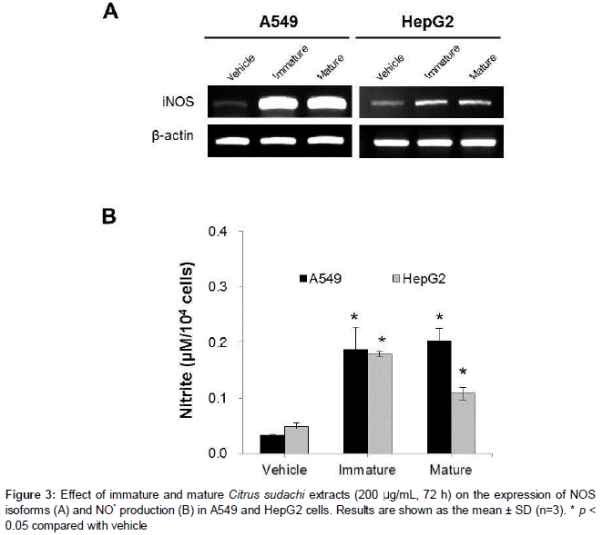
Results: Citrus sudachi exerted cytotoxicity in a time-dependent manner in cancer cells which increased with increase in maturity but did not affect normal cells. Citrus sudachi was found to induce accumulation of cells in the sub-G1 cell cycle phase, fragmentation of DNA and cell death with characteristics of apoptosis, in both types of cancer cells. Moreover, Citrus sudachi upregulated cellular NO• produced by activation of nitric oxide synthase (NOS), while it suppressed the levels of IAP mRNA in both types of cancer cells.
Conclusion: The results obtained suggest that Citrus sudachi induces apoptosis in A549 and HepG2 cells, which may be mediated by NO•. There is need for further studies on the role of Citrus sudachi in cancer treatment.
Introduction
Lung cancer is the leading cause of cancer death worldwide; it is responsible for 1.59 million deaths annually [1]. Liver cancer is the third leading cause of cancer death, causing 745,000 deaths per year as of 2012 [1]. Continuous efforts are being made to find novel, naturally occurring bioactive compounds that prevent lung and liver cancer because of the adverse effects that are often associated with artificial chemotherapeutic agents [1].
Citrus sudachi Hort. ex Shirai (Rutaceae), or, simply, sudachi, is primarily used as a seasoning in savory foods and as a flavoring in alcoholic beverages in Japanese cuisine but also used in other Asian food. Citrus sudachi is harvested before the fruit is fully ripe, while still green, because the flavor and odor are strongest at that stage. As the green color disappears, the acceptance of the fruit by consumers is gradually reduced. When fully ripe, the fruit turns yellowish, but it is generally used before that stage.
Citrus sudachi contains high levels of polyphenols and flavonoids [2-4]. Flavonoids extracted from the peel of Citrus sudachi have been shown to exhibit several biological activities that are beneficial to health including antioxidant, anti-inflammatory, antimicrobial, and antidiabetic properties [5-9]. These beneficial properties suggest new uses for Citrus sudachi as a nutraceutical, and studies of the immature and mature fruits of Citrus sudachi are currently of considerable interest in the pharmaceutical and food industries. However, the anticancer effects of Citrus sudachi have never been investigated.
The purpose of this study was to investigate the cytotoxic effects of Citrus sudachi harvested at two stages of maturity (immature and mature) on two tumor cell lines, human lung adenocarcinoma and hepatocellular carcinoma cells. This study demonstrate that nitric oxide (NO•) and inhibitors of apoptosis (IAPs) is necessary for the anti-tumoral capacity of Citrus sudachi, thus providing a strong basis for understanding the molecular mechanisms of apoptosis in A549 and HepG2 cells.
Methods
Plant material and extraction
Immature and mature Citrus sudachi fruits were harvested in September and November of 2014, respectively, on Jeju Island, Korea. The Citrus sudachi was mostly grown in Korea. No specific permissions are required for getting in Korea. Citrus sudachi was identified by Professor Jung Hyun Kim from Cheju Tourism College. Immature and mature Citrus sudachi fruits were then thoroughly rinse with tap and distilled water and were air-dried at room temperature. The air-dried fruits were grounded into powder using a blender.
Dried powder (5 g) of Citrus sudachi was extracted with 100 ml of 100 % methanol at 25 ºC for 72 h with constant shaking. The extract was then purified using a Sep-Pak C18 cartridge and a 0.45 μm membrane filter (Waters, Milford, MA, US), concentrated using a rotary evaporator (Buchi Rotavapor R-200, New Castle, DE, US), freeze dried and finally stored at -20 ºC until further use. Dried extracts were reconstituted in dimethyl sulfoxide (DMSO, Amresco, Solon, Ohio, USA) for cell treatment.
Cell culture
Human lung cancer (A549), normal lung (16HBE140-) cells, human liver cancer (HepG2) and normal liver (CHANG) cells were kindly provided by Dr. M.H. Cho (Seoul National University, Korea). A549 and 16HBE140 cells were grown in HAM’s F-12 and DMEM/F-12 media, respectively. HepG2 and CHANG cells were maintained in MEM and DMEM media, respectively. The cells were cultured in the appropriate media containing 10 % fetal bovine serum (FBS), 1 % penicillin/streptomycin, and 1 % L-glutamine at 37 ºC and 5 % CO2.
Cell viability assay
Cell viability was estimated using trypan blue as described previously [10]. Cells were seeded in 100 mm cell culture dishes at 2 × 107 cells/10 ml. The cultured cells were treated with fresh medium containing various concentrations (0-200 μg/ml) of the Citrus sudachi extracts dissolved in dimethyl sulfoxide (Amresco, Solon, Ohio, USA). DMSO alone was added to another set of cells as a solvent control (DMSO < 0.1 %).
The cells were then incubated for 24, 48, or 72 h, detached with trypsin, and then an equal volume of 0.4 % trypan blue solution (Lonza, Walkersville, MD, US) was added. After 1 min incubation, viable cells were counted with a hematocytometer. All experiments were done in triplicate.
Cell cycle analysis
Flow cytometric analysis was performed as previously described [11]. Cells (2 × 107 cells/10 ml) were treated with Citrus sudachi extracts for 72 h. They were then harvested and washed twice with 1 ml PBS, then centrifuged at 1000 rpm for 10 min at 4 ºC. Pellets were fixed with 70 % ethanol and kept on ice for 30 min. The cells were stained with 500 μg/ml of propidium iodide (Sigma, Louis, MO, USA) on ice, digested with 10 μg/ml of ribonuclease A (Amresco) at 37 ºC for 30 min, and analyzed with a BD FACS caliburTM flow cytometer (BD biosciences, San Jose, CA, US). The percentage of cells with DNA content less than 2N (sub-G1 phase cells) was analyzed with CELL QUEST software.
DNA fragmentation
A GeneluteTM mammalian genomic DNA miniprep kit (Sigma) was used to perform a DNA laddering assay [10]. To detect the DNA ladder, the extracted DNA samples were run on a 1.8 % agarose gel in TBE buffer. After electrophoresis, the gels were stained with ethidium bromide (EtBr, Sigma) and visualized with a UV light transilluminator.
Determination of nitrite production
The amount of NO• in the culture medium was measured by the Griess reaction as previously described [11]. Furthermore, 100 μl of cell culture media was mixed with an equal volume of Griess reagent and incubated at room temperature for 10 min.
Absorbance at 540 nm was measured using a Packard EL430 microplate reader (Bio-Tek Instruments, Winooski, VT, US), with fresh culture media serving as the blank. Results are expressed in μmoles per 104 viable cells.
Reverse transcription-polymerase chain reaction (RT-PCR) analysis
Total cellular RNA was isolated from each cell line using TRI reagent (Sigma) and semiquantitative RT-PCR analysis was carried out using a TOP scriptTM one-step RT-PCR kit (Enzynomics, Daejeon, Korea) [11]. Primer sequences were as follows: iNOS, sense 5ʹ-CCAGTGACACAGGATGACCTTCAG-3ʹ and antisense 5ʹ-TGCCATTGTTGGTGGAGTAACG-3¬ʹ; survivin, sense 5ʹ-GCATGGGTGCCCCGA CGTTG-3¬ʹ and antisense 5ʹ-GCTCCGGCCAG AGGCCTCAA-3¬ʹ; XIAP, sense 5ʹ-ACACCATATACCCGAGGA AC-3¬ʹ and antisense 5ʹ-CTTGCATACTGTCTTTCTGAGC-3¬ʹ; cIAP-1, sense 5ʹ-AAGTTCCTACCCCT GTCCAATG-3¬ʹ and antisense 5ʹ-CAAGTAGATGAGGGTAACTGGC-3¬ʹ; cIAP-2, sense 5ʹ-CCTGTGGTTAAATCTGCCTTG-3¬ʹ and antisense 5ʹ-CAATTCGGCACCATAACTCT G-3¬ʹ; and β-actin, sense 5′-GGTCATCTTC TCGCGGTTGGCCTTGGGGT-3′ and antisense 5′-CCCCAGGCACCAGGGCGTGAT-3′. 2 μg of total RNA was used in each reaction and was amplified in the exponential phase of PCR. Reactions were normalized to actin levels. Amplified DNA was electrophoresed on 1.5 % agarose gels and visualized by EtBr staining.
Statistical analysis
Data are presented as mean ± SD. Statistical analysis was done using a Student’s t-test (SPSS for Windows, 12.0, SPSS Inc. Chicago, IL, USA). Differences were considered significant at p < 0.05.
Results
Loss of cell viability
In this study, we examined the cytotoxicity of Citrus sudachi on A549 and HepG2 cells. Cells treated with 0, 50, 100, and 200 μg/ml of Citrus sudachi harvested at two stages of maturity (immature and mature) for 24 h responded similarly, with respect to viability, in that treatment with extracts from immature and mature fruits reduced the percentage of viable cells dose-dependently (data not shown). We then determined the time course of responses to 200 μg/ml of immature and mature Citrus sudachi in cancerous and normal cells treated for 24, 48, or 72 h (). The human lung adenocarcinoma cell line (A549) was more susceptible to Citrus sudachi to cell death than hepatocellular carcinoma cell line (HepG2). Exposure to 200 μg/ml of immature Citrus sudachi for 72 h reduced the viability of A549 and HepG2 cells to 35 and 46 %, respectively, whereas comparable values were found after treatment with mature Citrus sudachi (32 and 55 %, respectively) (). The extracts were not cytotoxic to either of the normal cell lines, indicating the potential specificity of the Citrus sudachi extracts against lung and liver cancer cells ().
Apoptosis identified by flow cytometric analysis
The population of cells in the sub-G1 phase in cells treated with 200 μg/ml of immature Citrus sudachi increased from 3.6 % (control level) to 12.2 %, whereas treatment with extract from mature Citrus sudachi increased the proportion of A549 cells in cell cycle arrest to 19.9 % (). Furthermore, treatment with immature and mature Citrus sudachi extracts resulted in increase in the sub-G1 population to 10.1 and 9 % in HepG2 cells, respectively ().
Internucleosomal DNA fragmentation
Our results revealed fragmentation of DNA following treatment of A549 and HepG2 cells with Citrus sudachi extracts, indicating the presence of apoptosis, whereas DNA fragmentation was not seen in non-cancerous cells (C). Mature Citrus sudachi resulted in a significant increase in the intensity of DNA laddering when compared with immature Citrus sudachi (C). These findings indicate that Citrus sudachi can induce apoptosis-mediated cell death in A549 and HepG2 cells, but does not cause apoptotic cell death in normal cells.
Expression of NOS isoenzyme proteins and total NO• production
In order to determine the role of NO• in apoptosis induced by Citrus sudachi, expression of iNOS and nitrite (NO2-) production is determined in A549 and HepG2 cells (). Immature and mature Citrus sudachi markedly up-regulated iNOS in both types of cancer cells; the increase was more pronounced in A549 cells (A). Rates of NO• production were calculated from the concentration of NO2- in culture media following treatment for 72 h. Cells treated with either immature or mature Citrus sudachi extract alone showed 2- to 7-fold increases in NO• production, as compared with vehicle-treated controls (B).
Changes in mRNA of IAP family caused by Citrus sudachi
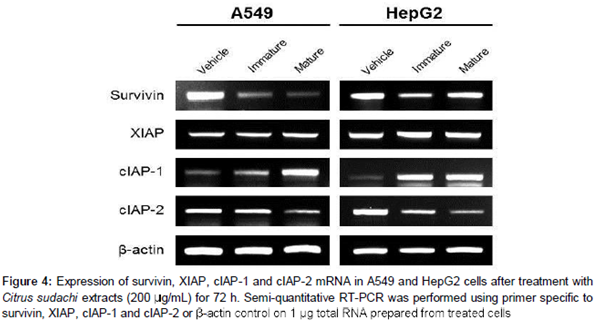
In this study, we investigated the mechanisms underlying cell death induced by Citrus sudachi, with a focus on the role of IAP family members (survivin, XIAP, cIAP-1, and cIAP-2). As shown , treatment with Citrus sudachi extracts for 72 h led to a reduction in survivin and cellular IAP-2 (cIAP-2) gene expression in both A549 and HepG2 cells. Meanwhile, the mRNA levels of cIAP-1 were enhanced by Citrus sudachi, but no differences were observed in mRNA levels of the X-chromosome-linked IAP (XIAP) ().
Discussion
Citrus sudachi is a plant with numerous health benefits including nutritional and medicinal advantages [5,8,9]. Medicinal plants are a common alternative to pharmacological cancer treatment in many countries. Cytotoxic screening of a number of plants has been done in order to investigate their anticancer activities and usefulness in drug development [12]. The aim of this work was to explore the biochemical and molecular events associated with programmed cell death in melanoma after treatment with Citrus sudachi harvested at two stages of maturity (immature and mature). Anticancer activities of Citrus fruit varieties have been found in other studies [13-16]. For instance, Citrus limon and Citrus paradise demonstrate cytotoxic activity in vitro against human prostate, lung, and breast cancer cell lines [13]. Qing Pi (Citri reticulatae viride pericarpium, immature tangerine peel) also strongly induced apoptosis in SNU-C4 human colon cancer cells [14]. The flavonoids extracted from Citrus fruits have been shown to inhibit the proliferation of A549 human lung adenocarcinoma cells [15,16]. Our results have clearly demonstrated that immature and mature Citrus sudachi inhibit cell growth and promote apoptosis in cell lines A549 and HepG2 (Figures 1 and 2). When developing anticancer agents from natural products, it is important not only to assess the selectivity (or specificity) in several types of cancer cells but also to assess if natural products exhibit any cytotoxicity in non-cancer cell lines [12]. Thus, the cytotoxic effects of immature and mature Citrus sudachi on the 16HBE140- (normal human bronchial epithelial cells) and CHANG (normal human liver cells) lines was tested. Given the fact that no cytotoxic effects were observed against normal cells, our data clearly indicates that Citrus sudachi exert a selective antiproliferative activity against lung and liver cancer cells (). Citrus sudachi was tested to evaluate whether this cytotoxic effect is associated with apoptosis. Induction of apoptosis in cancer cells has been used as an indicator of the ability of naturally-derived active components to treat cancer [17]. Fragmentation of DNA into 180-200 bp fractions is considered one of the hallmarks of apoptosis [18]. As shown in , DNA fragmentation as well as flow cytometric analysis showed that apoptosis was a major contributor to cell death induced in A549 and HepG2.
Data presented here of increased NO• production with Citrus sudachi treatment and the activation of iNOS level, suggest that NO• regulation accounts for the mechanism of Citrus sudachi’s action in these cell lines. NO• released by the inducible NOS (iNOS) is generated for long periods, by cells of the immune system among others, and has been shown to be a principle factor in inflammatory processes and cancer formation [19]. The role of NO• in cancer is multi-dimensional and depends on timing, location and concentration. High levels of NO• are cytotoxic to cancer cells [19]. The contrast, at low levels, constitutive production of endogenous NO• can promote tumor growth by inducing anti-apoptotic effects in many tumor types including melanoma [19-21]. Conflicting evidence exists regarding the ability of Citrus fruit varieties to modulate NO• production. While some investigators found that Citrus enhances iNOS expression and NO• production, others observed suppression. For example, Citrus polyphenol limonene promoted NO• production in several studies, leading to an inhibition in proliferation of human melanoma A375 cells [22], leukemia HL-60 cells [23], and epidermoid carcinoma A431 cells [24]. On the other hand, Sudachitin, a polymethoxyflavone from Citrus sudachi, inhibited LPS-mediated NO• production and iNOS protein expression in RAW264.7 macrophages [9], and Citrus nobiletin reduced NO• levels and decreased iNOS expression in hepatocytes [25]. Our data indicate a direct relationship between increased NO• production and loss of cell viability (Figs. 1 and 3) following treatment with both immature and mature Citrus sudachi extracts. To the best of our knowledge, there are no other reports on the effect of NO• stimulated by Citrus sudachi extracts on the proliferation of cancer cells.
The inhibitor of apoptosis (IAP) family of proteins are important regulators of apoptotic signaling cascades as they block the activity of caspases [26]. Therefore, IAPs are potential targets of Citrus sudachi. In the present study, immature and mature Citrus sudachi exposure suppressed survivin as well as cIAP-2 mRNA expression in both A549 and HepG2 cells. Survivin is a structurally and functionally unique IAP in that it contains a single BIR but no RING domain, and plays dual roles in mitosis and apoptosis [27]. cIAP-2 regulates not only caspases and apoptosis, but also inflammatory signaling and immunity, mitogenic kinase signaling, and cellular proliferation, as well as cell invasion and metastasis [28]. Notably, Citrus fruits reduce the anti-apoptotic signals of IAP proteins, thus facilitating the process of apoptosis in human melanoma A375 [21] and leukemia U937 [29] cells. Our results show for the first time that Citrus sudachi induces apoptosis in lung and liver cancer cells and revealed the role of the IAPs survivin and cIAP-2 in the process.
Conclusion
Citrus sudachi extracts induce apoptosis in A549 and HepG2 cells via NO• production and down regulation of IAPs, and that the effects depend on the growth stage of Citrus sudachi. This is the first report to reveal the possible mechanism of the antiproliferative effect of Citrus sudachi for the prevention of lung and liver cancer in cell culture model. However, further studies are needed to validate our finding in detailed in vivo studies.
Declarations
Acknowledgement
References
Archives


News Updates